 |






作者:John H. Lawton 来源:公众号生态学文献分享
摘要
定律(law)是指基于在特定条件下经常发生的一系列事件或过程的广义表述;具有广泛观察的趋势。基于此,我认为,生态学有许多定律,以自然界中广泛的、可重复的格局的形式出现,但几乎没有任何定律是普遍真实的。换言之,生态格局和支撑其的定律、规律和机制通常取决于相关的生物体和它们的环境。这种偶然性在相对简单的生态组织水平上是可控的(例如单个和少量物种的种群动态),并且在宏观物种集合、宏观的空间尺度或长的时间尺度内,以一种可控的形式重新出现,即最近称为“宏观生态学(macroecology)”的无细节(detail-free)统计格局。在中尺度上,偶然性变得异常复杂,这是群落生态学的特点,大量的案例除了进行薄弱的、模糊的归纳,几无可取之处。这些论点集中关注群落生态学的典型研究实例,并通过对比的方式,说明局域物种丰富度和区域物种库大小之间出现的宏观生态学关系。尽管在其产生过程中涉及大量的偶然过程和相互作用,但局域与区域丰富度图所说明的新兴格局是极其简单的。为了发现自然界的普遍格局、定律和规律,生态学可能需要减少对群落生态学“中间地带(middle ground)”的关注,减少对还原论和实验操作的依赖,而加大对宏观生态学的研究力度。
1. 引言和定义
在生态学中当然存在普遍定律。然而,几乎没有非常普适的定律。定律一词最恰当的含义应该是:“基于在特定条件下经常发生的一系列事件或过程的广义表述;具有广泛观察的趋势”。这个定义中没有说一个定律必须是普适的真实;只是说定律通常是真实的。
某些科学领域,例如物理学领域,有其深刻的、非常普遍的定律——这是生态学所羡慕的。生态学拥有一些普适定律的部分所有权,稍后会讨论。在这篇文章中,我特别想思考为什么生态学没有太多普适性定律。顺便说一句,生态学并不是唯一一个处于这种情况的学科。
有三个深层次的普适性定律支撑着所有的生态系统,一组一般的物理原理可以组合在一起构成第四条定律,还有一条定律如此简单,它实际上只是一个观察。它们是:
1. 热力学第一定律和第二定律。
2. 化学计量学的规律。
3. 达尔文自然选择定律。
4. 控制气体和液体的扩散和传输、皮肤和骨骼的机械特性、空气动力学和流体动力学等的一组普遍物理原理。
5. 微不足道但重要的观察是生物体彼此之间及其与环境的相互作用。
现在先来定义一些术语。格局是我们在自然界中观察到的规律性(regularities);即,它们是“广泛可见的趋势”。这些都是定律1-4和观察5的共同作用产生的;当我们试图理解生态学中的格局时,大量的努力集中在试图解开由定律3、4和5创造的戈尔迪乌姆之结。尽管这实际上与上面的词典定义不一致,但大多数生态学家认为定律和规律是支撑和创造格局的普遍原理,就像体育和其他游戏的定律(或规律)创造了人类活动的丰富格局一样。许多人认为,定律比规律更强大、更根本、更普遍。最有用的科学定律对自然界的作用有深刻的洞察力;规律不那么宏大。机制(Mechanism)则更弱;一个或多个机制可以产生独特的现象;但普遍规律需要共同的机制。归纳(Generalisation)是一个不精确的术语,它暗示着一些经常发生的事情,无论是格局还是规律。假设(Hypotheses)和理论(theories)是试图弄清楚定律或规律是什么;假设比理论更具试探性,极其成熟的理论最终会披上定律和规律的外衣。模型(Models)是将定律或规律以数学形式编纂的理论或假设。
在生态学中(以及以上1-5),我们根本的定律和规律并不是普适性的。因为环境不同,生物体相异,我们最终得到的定律、规律和机制也不同,即,它们是偶然的或情况依赖性的。“偶然的”的意思是“只有在特定或规定的情况下才是正确的”。一条偶然的规律(或定律)的形式是:如果A和B成立,那么X就会发生,但如果C和D成立,那么Y就是结果。由此得出的结论是,格局也将是偶然的,理论也是如此。从陨石的撞击,到变幻莫测的偶然性突变,一些偶发事件可能是最广泛意义上的“历史意外”。
请注意,相同的格局可以由不同的规律生成(格局并不意味着过程),而相同的基本规律(例如数学模型)可以生成不同的格局,这取决于偶然的细节。
在清理和润色了语义层之后,我们可以开始讨论这个问题了。我将论证,生态学有大量的(偶然的)格局,有定律和规律的支持,但要发现有用的归纳,我们需要看看真实的格局。最有用的偶然性理论和最醒目的偶然性格局都出现在相对简单的系统中(如种群),也出现在非常大尺度的系统中(例如物种-区域关系)。相比之下,群落生态是一团乱麻,偶然性如此之大,以至于很难找到有用的归纳。
2. 生物群系、气候、宏大图景、杂乱的细节和偶然性理论
《群落和生态系统》(Communities and Ecosystems)第二版(Whittaker 1975)第167页的图4.10显示了与两个简单环境变量-平均年降水量和年平均温度-有关的陆地世界生物群的主要类型。苔原、沙漠、温带森林、热带森林、稀树草原等都是这个简单环境空间中的特色部分。正如Whittaker指出的那样:“不同类型之间的边界不可能精确定位”,例如,气候并不是唯一决定生物群出现地点的因素;土壤和火也起到了作用。但作为一个很好的第一近似,支撑这一简单而醒目的自然界格局的机制已被充分理解,作为一种组织自然的方式,它与天文学家在尝试对恒星进行分类时所做的一切一样好。
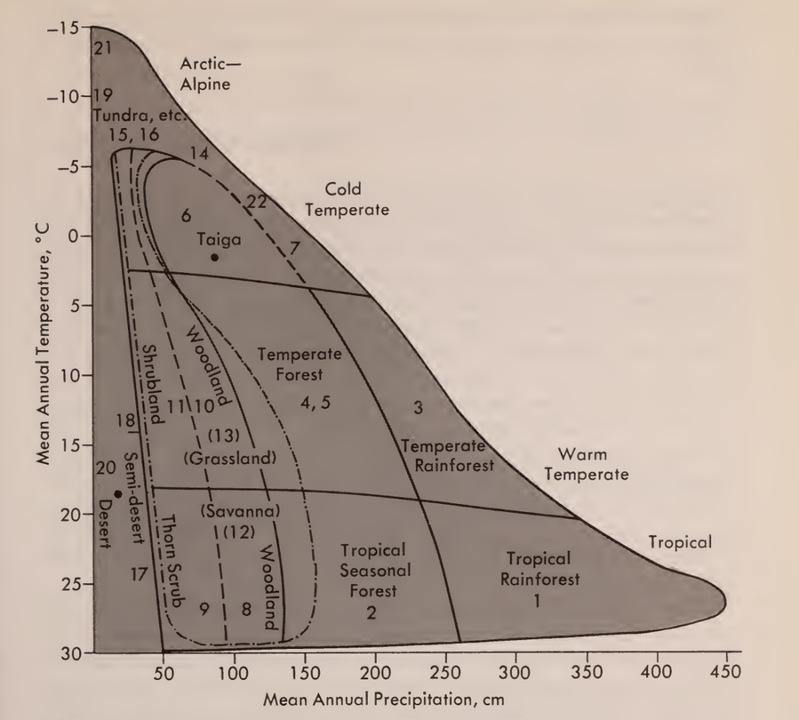
图1. 与气候湿度和温度相关的世界生物群落类型格局
Whittaker的图表所揭示的格局是醒目和明显的,因为他避免了陷入细节的泥潭。在局域范围内,在较小的尺度上,自然将创造无穷无尽的多样性,归纳消失,或者至少变得更难看到。但Whittaker认识到的格局不可避免地是连续的。降雨和温度是决定世界陆地生物群的主要因素,但它们的影响在一定程度上取决于土壤、火灾、其他气候细节(例如沿海和大陆的影响)和演化历史(桉树在澳大利亚打破了规律)。正是这种偶然性,事实上几乎不可能找到生态学中普适性的格局。这一点,加上许多生态学家有一种近乎自杀的倾向,他们以醒目的一阶现象为代价,忽视复杂性和细节。当然,细节很重要。但是看到树木之前,还是要先看到森林。
3. 种群动力学与偶然性理论
3.1. 综述
你认为现实世界中有多少种种群动态?每个物种都有不同的动态吗?地球上可能有一千万种动植物;真的有一千万种种群动态吗?
某种程度上,答案显然是“Yes”。事实上,它比“Yes”更糟糕,因为同一物种的不同种群也可能表现出明显不同的动态。在完全不同的层面上,答案是响亮的“No”。并不是有一千万种种群动态;相反,在几个共同的主题上,有许多本质上微不足道的变化。具体有多少还有待观察,但并不多。种群动力学理论的一个关键作用是阐明和定义这些类别的种群和集合种群行为,并指定可能导致每一类行为的生物条件、物种相互作用和环境(定律、规律和机制)的种类,例如稳定平衡、极限循环、混沌或持续但随机的波动。这里的关键偶然变量包括物种的内禀增长率和它们所处的食物网的复杂性。基于偶然性理论的第二类归纳,可能是预测不同种类的种群对中等或重大干扰的反应。
换言之,种群动力学的理论,对生态规律的探索,取决于有机体和它的环境。这种理论是否真正具有预测性是值得怀疑的。也就是说,在给定物种名称或有机体类型以及它在哪里生活的情况下,生态学家可以以任何程度的确定性来指定它将显示的种群动态的类型,而不必实际看到时间序列,也不知道关于其生物学的任何其他信息。但所有证据都表明,偶然性理论并不复杂和多维,以至于生物体所显示的种群动态范围完全无法通过一套定义明确的规律和机制来理解。
3.2. 将普遍的偶然性理论转化为管理工具
当规律和机制与种群管理者面临的日常实际问题纠缠在一起时,困难就开始了,这些人是处于最前沿的生态学家,他们必须控制害虫、维持渔业或拯救濒危物种。没那么深思熟虑的批评者指出,由于目前的理论模型不能为特定问题提供直接、实用的解决方案,生态学是一门软科学,更糟糕的是,该理论没有意义,因为它缺乏预测能力。坦率地说,就这些批评者而言,没有规律可以立即为紧迫的问题提供答案。
如果我们需要或想要详细预测特定物种在特定栖息地的种群动态,只能在感兴趣的地方或栖息地详细研究该物种,除此之外别无选择。通过利用类似物种(例如,具有相似身体大小和营养位置的分类近亲)的动态知识,可以在中级水平上取得一些进展,但详细的预测需要详细的知识。
顺便说一句,一些种群可能表现出混乱的动态,这显然意味着,就像天气预报一样,我们将无法预测未来很远的准确轨迹。但混沌波动在吸引子表面是有界线的,因此它们具有可预测的性质。
3.2.1. 具体的管理格局不是在理论真空中开发的
没有人在真空中构建详细的种群管理模型。他们使用相同的普遍原理来阐述和借鉴基本的(或有)理论模型。有一个特别好的例子是Potts(1986)开发的模型,该模型来自英格兰南部福丁布里奇的猎物保护信托基金,作为灰山鹑(Perdix perdix)的管理工具。灰山鹑是重要的猎鸟,或直到最近种群数量严重下降。Potts和他的团队开发了模型,将其作为一种管理工具,用于理解下降的原因,并设计出扭转下降的方法。该模型预测了稳定的、依赖于密度的反馈的普遍动态,这一点在数千种其他动物种群中都有体现。如前所述,真正独特的种群动态类别很少,而灰山鹑很容易被归入其中一类。但作为一种管理工具,该模型也表明,在同一主题上有大量次要的、尽管实际上很重要的变化。该模型成功地预测了英国不同耕作和捕食者控制制度下的不同种群水平,并了解了适当参数的变化后,似乎可以很好地预测欧洲大陆和北美地区这些鸟类被引入的山鹑数量。它是一种很好的管理工具,基于优秀的应用科学。
然而,我们清醒地认识到,无论我们的偶然性理论变得多么好,几乎不可避免的是,具体的种群管理问题将通过针对具体地点的研究来可靠地解决,以便为手头的任务设置参数和完善通用模型。在现实中,这意味着种群管理问题的好答案不太可能是快速答案。有时,在决定性时刻,除了快速而经干预的答案之外,可能别无选择;基于健全理论的快速而经干预的答案,胜过胡乱猜测或不知情的猜测,或者根本没有答案。无论如何,只要有时间,生态学家必须要求重新提供来源以提供经过适当研究的答案。这样做并不意味着每一个案例都是如此特殊的,以至于没有规律。这些规律是偶然的,对于真实的种群来说,它们可能很难制定。
4. 群落
4.1. 群落在数量级上更为复杂
我认为,生态学中没有规律或定律,主要是因为许多从业者过于强调“群落生态学”-即在局域尺度上相互作用的共存物种集合的生态-以及同样强烈地强调进行控制性野外实验,以发现这些组合是如何构成的。不管是否有一些细节,对于生态学的这一领域,人们的看法是正确的。令人痛苦的是,很少有模糊的归纳,更不用说规律或定律了,而且必要的偶然性理论看起来也很复杂。让我花一些时间来为这一挑衅性的声明提供背景。
在接下来的文章中,我用“群落(community)”这个术语非常松散地指相互作用的物种的大集合,并尽可能地遵循Fauth等人(1996)的说法使用了更准确的术语“局域种团(local guild)”和“组合”。
4.1.1. 蕨类昆虫和其他系统
二十年来,我研究了英格兰北部约克郡Skipwith Common的一片蕨类植物-以蕨类植物为食的局域昆虫种团。在研究期间,这片蕨类植物每年平均有17种以上,最少有15种,最多有19种。我对局域这个小小的昆虫种团结构了解了很多。大多数物种种群是由密度依赖过程调节的,它们的丰度受到自上而下和自下而上力量的混合影响。其结果是在物种丰度的等级顺序上具有高度的时间可预测性(稀有物种保持稀有,常见物种保持常见),尽管这种可预测性随着时间的推移而缓慢下降(等级丰度两年比二十年间隔更接近)。系统中物种的种类(它们如何和何时进食),而不是它们的丰度,似乎受到了避免蚂蚁捕食需要的强烈限制。种间竞争起不到明显的作用。
作为一名群落生态学的研究人员,我在Skipwith Bracken斑块上采取的方法,以及我获得的各种见解,都大致符合这门课程的要求。在所有这些和许多其他案例(包括我的案例)中,目的都是寻找“组装规律(assembly rules)”;试图理解为什么一些物种是常见的,另一些是稀有的,为什么有些物种被排除在外;试图厘清竞争和捕食、自上而下和自下而上的力量、或对组成种群的干扰的相对重要性,等等。
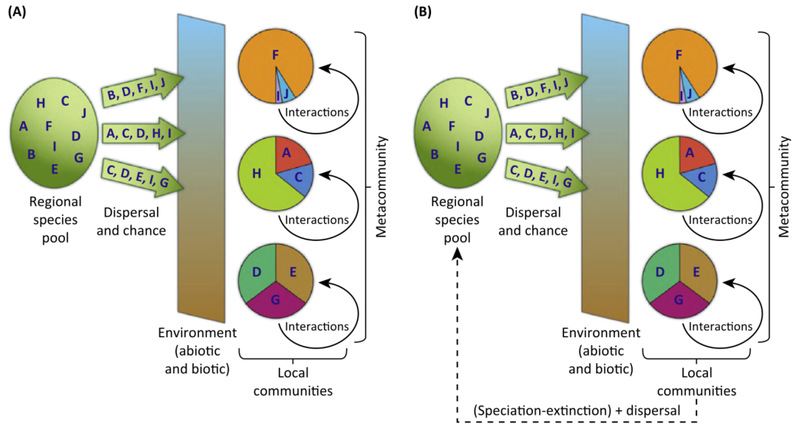
图2. 群落组装的两种模型
这些见解来之不易,尽管存在挑战,生态学在建立一系列关于局域群落组装的案例方面取得了长足的进步。基本结论是,重要的过程和由此产生的群落动态在不同的系统之间往往是显著不同的。群落生态学的理论取决于所涉及的有机体和它们的环境。这些规律在很多方面都是偶然的(我们稍后会看到),以至于寻找格局是行不通的。这是来之不易的洞察,我们不能掉以轻心。矛盾的是,它实际上是一个有用的归纳!自然界不需要这样的组织-所有局域物种的组合都可以按照完全相同的规律工作。
4.1.2. 太多偶然性
尽管现在已经很好地了解了自然界中几组相互作用的局域物种是如何工作的,但问题是无法预测哪些过程在哪些系统中是重要的。从这个意义上说,研究群落与研究种群动态没有什么不同。
处理众多特定实例的一种方法是对它们进行分类,以辨别新出现的格局及其基础规律,即识别和定义偶然性。Schoener(1986)提供了迄今为止最全面的尝试,为群落生态学发展一套偶然性理论,不是基于分类学,而是基于他所说的有机体生物学的两个“原始轴(primitive axes)”和环境。这是一项英勇的努力,将群落划分为不同的类型,从而找出不同生物群和栖息地的相互作用规律。
Schoener定义了六个原始生物轴(身体大小、(自然种群)增长(recruitment)方式、世代时间(generation time)等)和六个环境轴(物理因素的剧烈程度、资源投入的性质、空间破碎化等)。由于各种有机体和各种环境之间没有相关性,那么就有12个关键驱动因素创造了基本模板,以及在这个n维空间的特定点上的大量独特组合,在这些组合中,物种可以进行群落组装的游戏。难怪构建群落的过程在不同的系统之间有明显的不同。
Schoener在书的最后一章表示,他对我们将能够通过沿着他所倡导的路线发展出一种强大的、偶然的群落生态理论持谨慎乐观的态度。我不太确定。他提醒人们注意对群落结构施加的或有约束的复杂性,这是完全正确的。正如我已经指出的,生态学家应该庆祝我们在理解这一点方面取得了多大的进展。但我怀疑,努力将每一项研究都局限于由至少12个原始轴组合定义n维空间中的一个点,是否值得。自从Schoener的模型发表以来的十年里,没有人认真地尝试使用和测试它,这表明它可能太复杂、太难了,以至于没有用。
基本信息是令人深省的。通过对特定系统的精心研究,我们可以广泛地理解在一个地点和时间相互作用的物种的局域规律。但撒旦存在于偶然的细节中。几乎每个地点、时间和物种的组合都足够不同,以至于几乎不可能找到更多的普遍格局和规律,即使有时我们可以找到薄弱的归纳,我们也永远无法根据对6个原始有机体轴和6个原始环境轴(或任何其他必然同样复杂的偶发事件集)的先验知识来预测它们。与种群动力学不同,我怀疑我们是否能为管理目的建立一个有用的、实用的模型,即使是10或20个物种组合(更不用说数百个物种了)。当然,另一种观点是,我(和其他人)只是缺乏尝试的想象力和勇气。
4.1.3. 整个湖泊的操纵
有一个领域的观点显然是错误的,那就是湖泊学。整个湖泊操纵的科学(例如,在食物链的顶端添加或移除捕食性鱼类)是生态科学的成就之一。当然,这里有一些相对简单的规律,这些规律针对的是数量可控的变量。若非如此,这种湖泊管理技术就不会奏效。当然,湖泊是复杂的生态系统,有着复杂的生物群落,涉及数百个物种?是的,但就手头的任务而言,它们并非如此。
湖泊中的营养级联涉及相对较少的关键生物,并且是简单的近乎线性的食物链(食鱼性鱼→食浮游生物鱼类→浮游动物→浮游植物)。该湖的大部分多样性位于其他地方,在底栖生物和边缘的芦苇丛中。因此,这个问题基本上是种群动态的问题,而不是群落生态学的问题。即使在这个相对简单的系统中也有重要的偶然性事件,但不会太多以至于变得不可行。类似的评论也适用于巴哈马群岛上的试验性食物链(蜥蜴→蜘蛛→昆虫→植物)。这可能是目前理论生态学所能达到的最复杂和最偶然的情况。它清楚地表明,存在普遍的格局,以及支撑这些格局的有用规律;但它也划定了当前进展的边界。
4.1.4. 其他类型的归纳
在继续之前,值得指出的是,在群落生态学中,从案例的的翻箱倒柜中,涌现出了性质完全不同的普遍性结论。例如,我们现在知道通过食物网传播的几种“间接相互作用”在确定局域群落成员资格和种群丰度方面发挥了重要作用,令人惊讶的是,在群落的实验操作中,间接相互作用并不比直接相互作用花费更长的时间来揭示其本身。这当然不是一条定律,但它是一个有用的归纳。例如,它警告我们,当群落以某种方式被操纵或改变时,要预料到它们会发生意想不到的变化;间接相互作用会给你惊喜。
另一组缓慢推进、后退、重组甚至可能再次前进的归纳是食物网中的格局。十年前,它看起来确实像是食物网中有重复的格局,并有一个理论框架来解释它们。现在,由于有了更好的经验数据,整个事情看起来更加不确定。我仍然认为,食物网的结构是有规律可循的,但我必须承认,我可能错了。
4.1.5. 试图找到群落生态规律的其他问题
总而言之,生态学家非常正确地认为,仍在进行中的群落生态学方面的巨大研究努力并没有产生太多的格局或可行的偶然规律。这种情况还有其他原因。在这里将它们与我已经触及的一些问题简单地结合起来是很有用的,从而将论点的实质塞进几个短段落中:
1. 对于大多数多物种组合,可以花很大力气弄清楚系统如何组合的一些细节,但每个系统似乎都不同,无法预测一个新的、未经研究的系统的局域规律是什么。
2. 在一个地方和时间对物种的相互作用有了来之不易的认识,人们担心重要的过程会随着时间的推移而改变。根据定义,它们必然会在演替系统中发生变化,从长远来看,红色的环境噪声将摧毁所有自然界的局域结果。但如果局域规律的衔接是暂时的,那么可能很难用它们来做出预测。
3. 在一个地方以巨大的代价学到的关于群落内物种相互作用的知识,也可能不容易转移到“其他地方”大致类似的系统中。更有可能,在有限时间内,在一个地方存在关键的相互作用;但同一物种在不同的地方可能会产生更微弱的影响。
4. 间接效应的存在使得即使对研究很透彻的系统进行预测也很困难。
5. 矛盾的是,也许传统的群落生态学的主要弱点,以及为什么它未能提出许多格局、规律和可行的偶然理论,是它过分强调局域性。
总而言之,群落生态可能是世界上最糟糕的。它比种群动力学更复杂,所以偶然性理论不起作用,或者更确切地说,偶然性本身太复杂而没有用处。但矛盾的是,群落生态并不够大,也不够醒目,不足以打破它似乎深陷其中的压倒性复杂性。所有这一切都回避了一个问题,即为什么生态学家继续将如此多的时间和精力投入到群落生态学的传统研究中。在我看来,向前看的时候到了。一个很好的解决方案很可能是进一步扩大努力的尺度,达到这样的程度,所有特殊的细节都会被“平均掉(average out)”,生活再次围绕一些相对简单的基本规则组织起来。Whittaker的生物群系就是一个典型的例子。
5. 宏观生态学
5.1. 综述
宏观生态学是生态学、生物地理学和演化论的混合体,它试图超越局域群落组装令人费解的细节,找到一个更大的图景,从而使一种统计学秩序从混乱中出现。Jim Brown和Brian Maurer将宏观生态学确定为一个新兴的生态学学科,并为其命名,为我们的科学带来了巨大的帮助。
迄今为止,这门新兴学科的主要重点是在物种水平上的格局,但我认为没有任何逻辑上的理由,为什么这种方法不能包括生物组织的更高单位,从Whittaker对生物群落分布的分析,到生态系统结构和功能的主要格局和规律性。
人们已经做了几十年的宏观生态学,但没有给它一个专门的名字。种群丰度的的典型对数正态分布、岛屿生物地理学理论、热带-温带梯度,等。原则上,这些现象中的每一个都可以通过对所有涉及的物种进行详细的、还原主义的研究,通过研究它们的生活史、天敌、竞争者、其他相互作用、扩散率等等来揭示。但实际上,这样说来,这项任务显然是无望的!只有忽略细节,才会出现格局。
5.2. 局域物种丰富度与区域物种库尺度之间的关系
为了了解我们的方向,请考虑一个最简单的问题,我们可以对一个局域种团或一个组合提出问题。是什么决定了它所包含的物种数量?如前所述,我观察到每年平均有17种食草昆虫在Skipwith的蕨类植物上觅食。为什么是17种?为什么不是2种,或170种?令人惊讶的是,群落结构的所有方面中最基本的一点可能与主导群落生态学中许多传统思维的局域过程几乎没有关系。这个问题也不一定能通过做任何数量的小尺度操纵性实验来回答。为了回答这个问题,我们需要采取一种宏观生态学的方法。
5.2.1. 主要的环境过滤器
群落组装、局域种团等,是一个多阶段、多层次的过程,它始于一个区域的物种库。这个物种库存在于一个生物地理区域内,其空间尺度比局域群落的空间尺度要大很多倍。理解物种库的起源需要了解生物群的演化史、地质学、板块构造等方面的知识。大多数生态学家只是简单地接受了物种库的存在,而忽略了其重要性。Ricklefs有力地论证了这种短视是不明智的。
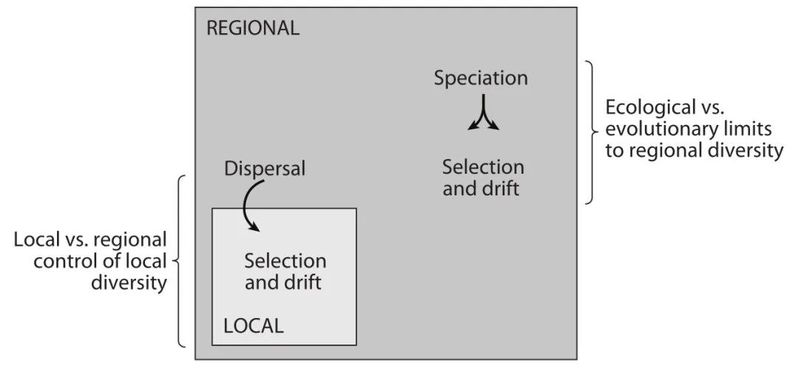
图3. 生态群落的四个过程框架来说明关于对局域和区域物种多样性的主控关系
局域群落通过一系列的过滤器或阶段从这个物种库中组装起来。这些过滤器本身在不同的空间和时间尺度上工作,并有重叠。但大致上,它们是这样的。首先,物种必须到达并建立种群。距离,与物种库的隔离,排除了一大部分物种。其他物种到了那里却发现栖息地不合适。强大的环境过滤器对所有的群落都起作用。更加微妙的过程在景观尺度上运作,介于区域物种库和局域群落之间-Holt(1993)称之为“中尺度的生态学(ecology at the mesoscale)”。我们都熟悉物种-面积的关系,但除了面积的影响外,生境斑块的数量、形状和空间排列的不同也意味着种群持续存在和局域灭绝之间的区别,并将进一步过滤和塑造局域组合。只有在最底层,传统群落生态学的核心问题才会发挥作用-物种相互作用在筛选出通过所有其他大过滤器的幸存者中的作用。以这种方式来看,传统的群落生态学实际上是关于二阶,甚至可能是三阶的过程。
5.2.2. 第一类和第二类系统
考虑区域物种丰富度(物种库)和局域物种丰富度(群落、局域种团、组合等)之间的关系。在X轴上画出区域物种库的丰富度,在Y轴上画出适合于每个物种库的局域物种库的丰富度。由于每个物种不可能生活在所有地方,而且除了少数明显的例外,我们不期望区域物种库中的每个物种都出现在每个群落中。区域和局域物种丰富度之间关系的最简单的模型是“(成比例的采样)”(第一类),局域丰富度与区域丰富度成正比,但低于区域丰富度。本质上,这个模型对局域物种相互作用的了解,不足以理解局域物种丰富度。局域的丰富度与区域物种库的大小成正比。
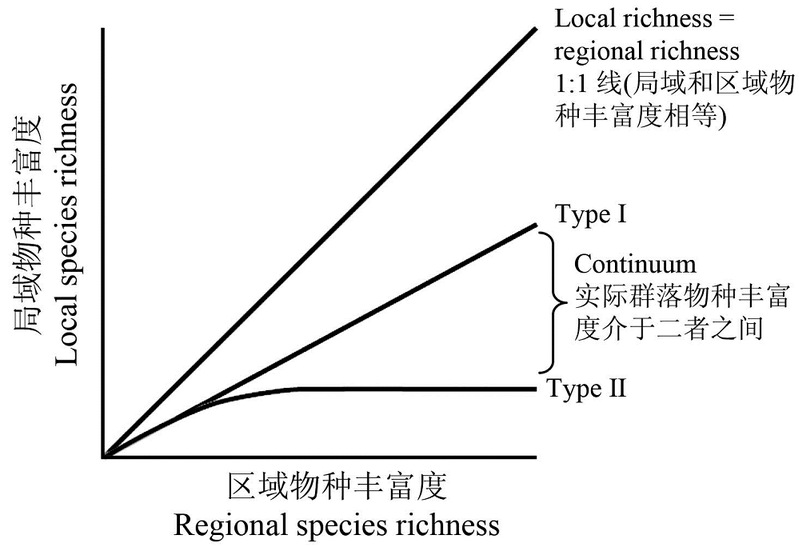
图4. 局域和区域多样性之间的理论关系
另一方面,如种间互作对群落构建很重要,那么(在其他条件相同的情况下)局域与区域丰富度的关系图将是一条负加速的曲线,达到一个顶峰(第二类)。从生物学上讲,从一个区域物种库中组装的相互作用的物种模型会产生很好的饱和曲线。真实的物种集可能位于一类和二类系统之间的任何地方,因为种间互作在限制局域物种丰富度方面变得越来越重要。但请注意,在这个层面上,可能的结果已经简化。我们不再被细节所淹没。理论只定义了两个极端的结果,加上中间的结果。格局再次出现。
不可避免地,有一些(偶然的)理论上的复杂情况。适度的环境干扰,在群落中淘汰一些物种,可以产生线性(第一类)的局域-区域丰富度图,即使在没有干扰的情况下,种间竞争强烈到足以产生第二类曲线。另外,模拟从区域物种库中取样的模型,其丰度符合广泛观察到的典型对数正态分布,可以产生“伪饱和”的第二类曲线,取样效率低,但完全没有物种间的相互作用。换言之,格局本身并不能定义机制。但无论基础机制是什么,真正的生态组合位于从第一类到第二类系统的连续体上,这个问题对群落生态学以及寻找自然界的格局和规律有重大影响。
如果第二类系统是常见的,由种间互作对局域的丰富度有严格和一致的限制,那么传统的群落生态学的关注就得到了证明;局域的过程比区域的过程更有优势。在这种情况下,群落生态学的议程侧重于小尺度的局域操纵实验,旨在解开群落组装的规律,这将是完全合理的,我们将有一个漫长的过程。但是,如果第一类系统,或弱曲线的中间系统是常态,这就不再正确了。为了了解一个局域组合的最基本的事实-它包含多少物种,以及由此必须产生的一切-群落生态学的传统议程看起来不必要地复杂。
5.2.3. 大多数真实的系统是第一类还是第二类?技术问题
根据目前的证据,对“大多数真实系统是第一类还是第二类?”这个问题的答案是“在大多数情况下,它们是第一类”。我的一个学生,Diane Srivastava,对文献做了全面的综述;这个总结在很大程度上借鉴了她的论文。首先,需要思考一些技术问题。
Srivastava指出,在文献中,对于适合于测试真实系统在这个模型框架中位置的数据类型存在混乱。例如,将同一地区的不同生境混在一起的数据,涉及不同类型的物种,尽管是广泛相关的物种,通常是不合适的,因为它们很容易产生人为的第一类关系。这些细节在这里并不重要,但意味着我们必须关注涉及类似分类群和栖息地的数据,这些数据来自不同的地区,或者(对于寄生虫和以植物为食的昆虫)同一地区的不同宿主物种。
Srivastava的第二点是,区分第一类(线性)和第二类(曲线、饱和)关系的正式统计测试远非简单的;例如,局域和区域物种丰富度在统计上并不独立,而且局域丰富度的数据可能是假重复(pseudoreplicated)的。在估计区域物种库的构成方面的困难和错误也会产生问题,并造成人为的关系(例如,高估物种物种库的大小,会产生错误的二类曲线)。
这类困难并不是局域和区域丰富度之间关系的唯一问题。在新兴的宏观生态学也遇到了类似的问题,这些数据很难获得,而且往往是为了手头的工作以外的目的而收集的。最好的前进道路不是被技术细节所掣肘,而是在两个方面谨慎地进行。首先是询问现有的数据究竟是什么样子的,尽管存在已知的问题,并得出初步结论-世界似乎是什么样子的暂定模型(tentative models)。其次是为任何一个似乎最能描述特定系统的模型寻求独立的证据。在目前的情况下,我们可能会在似乎是第二类的系统中寻求强烈的物种相互作用的实验或统计证据(对资源的种间竞争,通过共同的天敌的似然竞争,等等)。相反的预测显然是,在一类系统和弱曲线型、中间程度系统中,将不存在同等的强相互作用;或者(更微妙地)它们的影响被压倒性的过程所减轻,或者被物种分布中强烈的空间异质性所减轻。
5.2.4. 对例子的综述
大多数的生态系统似乎是第一类型的。如果只计算一次有一个以上分类单元数据的论文,目前至少有23个例子。其他研究也提出了一类关系,但所提供的数据不够完整,无法确定。
相比之下,有15篇论文描述了曲线型的关系。其中只有一些似乎是严格意义上的二类,饱和曲线;其余的是中等程度(intermediate)和弱的曲线型。对那些我们有理由相信关系是饱和的[强],或弱曲线型的[弱]的情况进行分类(其余的数据点太少,或有其他不确定的联系,无法确定),非线性的例子包括深海腹足动物(弱)、鱼类和两栖动物的寄生虫(强), 等。
并非所有这些例子都是由其原作者以我在此提出的方式绘制和分析的。如果结果可以根据这个模型进行合理的解释,或者对数据的重新分析产生了线性或曲线拟合,它们就被包括在内。也有少数情况下,原作者、Srivastava以及Cornell和Karlson得出不同的结论。例如,Hawkins和Compton(1992)发现曲线模型比无花果黄蜂的线性模型的拟合度稍好,但指出改进的拟合度是在局域-区域多样性图的“错误端(wrong end)”(即低多样性结束)。Srivastava将这个例子坚定地归入第一类,因为在中度到高度多样性系统中没有证据表明存在曲线性。Cornell和Karlson把这个例子列入他们的弱曲线性类别。
这篇论文的头数(head-count)并不具有任何定义性。其中许多论文存在一个或多个前面概述的统计学和方法学问题,而一些支持第二类模型的研究仅基于两个数据点,尽管区域物种库大小有很大差异,但却显示出相同的局域丰富度。这些研究被包括在这里,并以其表面价值为依据,但我们不能排除这样的可能性,即它们所代表的系统实际上是第一类,有两个随机抽取的点,恰好具有相同的局域丰富度。
考虑到所有这些注意事项,暂时的结论是,一类系统和弱曲线型的中间系统在现实世界中被报道的频率至少是饱和二类系统的两倍。事实上,如果我们简单地把这些数字加起来,并忽略“不知道(don't knows)”,一类和弱曲率系统可能比二类系统常见四倍左右。如果你愿意,我们可以把这当作我们目前的工作假设。考虑到群落生态学的所有复杂问题,出现的是一个相当简单的画面-出现的格局和规律,为生态学家可能想要回答的一个最基本的问题设定了两个可能的极端行为的界限:什么决定了我所选择的研究系统中的物种数量?通过超越细节,宏观生态学简化了我们需要知道的东西,并测量和揭示了世界上所有小尺度的、详细的、还原主义的实验所不能揭示的东西。
5.2.5. 这些结论是否得到其他证据的支持?
正如Cornell和Karlson(1997)所指出的,这些数据在一定程度上与人们在其他方面的预期大致一致。例如,在以蕨类植物为食的昆虫中,种间竞争通常不是一个强大或普遍的力量,也不是在其他局域的植食性昆虫种团中,这与现有的关于这个群体的研究都符合第一类的事实一致。但也有一些困惑,提醒我们类似的格局可以由一个以上的过程产生。两栖动物和鱼类上的寄生虫组合似乎有很强的饱和度,然而至少有一位研究鱼类寄生虫的专家认为,这些系统离物种的饱和度非常遥远。在数据中也很难看到许多其他分类学或栖息地相关的格局,这与前面讨论的试图识别和分类不同类型群落的困难是一致的。一些相同的分类群出现在两个阵营中,但当这似乎与生境有关时,却完全不清楚原因。因此,我们又陷入了没有任何先验的方法来知道一个潜在的研究系统是第一类,第二类,还是介于两者之间的问题。
5.2.6. 结论
在大多数研究中,局域物种丰富度的主要驱动因素似乎是区域物种库的大小,局域丰富度与区域丰富度呈线性或弱曲线关系。与局域丰富度的硬性限制相一致的数据是存在的,但它们不是大多数。局域丰富度对区域丰富度的强烈依赖可能是由于物种间的相互作用-传统群落生态学的东西-很弱;或者物种间的相互作用很强,但并不限制物种丰富度(例如特化种敌人强烈“自上而下”的影响);或者它们很强,但其影响被其他过程所取代,或空间异质性。这些规律是偶然的,但并没有偶然到无法理解的程度。不管是什么偶然的规律,在所有这些情况下,要回答关于一个群落的最基本问题-它包含多少物种,为什么?- 群落生态学家必须把他们的头抬到栏杆之上,成为宏观生态学家。当你这样做时,问题会改变,但格局会变得更简单、更清晰,并有可能发现一些简单的、尽管仍是偶然的基本规律。
6.其他宏观生态格局和结论
宏观生态学不是寻找生态学格局和规律的灵丹妙药。但就像温度、降雨量和生物群落之间的关系,或者局域物种丰富度和区域物种库的大小之间的关系一样,还有足够的其他例子让我相信,站在后面去了解更大的图景,将比目前困扰我们的主题的拱形还原论揭示出更广泛的生态格局和规律。格局和规律将是有条件的,我希望我已经说得很清楚了。它们不会成为非常普遍的定律。但是,根据目前的证据,在宏观生态学范围内出现了可控制的偶然性。
我不打算列出当前宏观生态格局的详尽清单,也不打算列出针对它们的一些标准。所有的格局都允许有一些例外,因为驱动过程是偶然的。但与少数几个相比,这些归纳性的东西要多得多,也更有说服力,而且从传统群落生态学中得出的结论往往很弱。对于一些人来说,有一些关于细节的术语,但这些论点主要是关于二阶现象,尽管这些现象对于理解基本过程可能很有趣和重要。
我特意选择了一系列不同类型的例子,包括一些生态系统级别的格局,以说明该方法的广度,并强调了一些明显的优点和缺点。其中一些格局将为几乎每个人所熟悉。我提出它们是为了证明,生态学中有大量的格局,有经过充分研究的规律和机制支撑,尽管广泛的格局不必在任何地方由相同的过程产生;可以说,最可靠的格局是那些有几种不同的理论解释的格局,因此可以由几种不同的机制产生(下面的4-7提供了很好的例子)。这些格局是:
1. 在动物群内的物种丰度频率中,有截断的、大致的对数正态分布,在物种地理范围的大小中,也有大致的对数正态频率分布。
2. 局域和全球不同体型的动物物种数量的截断对数正态格局 。
3. 物种丰富度、种群丰度、地理分布范围大小和体型之间存在有趣的三向和四向关系。
4. 物种-面积关系,主要是(但不总是)双对数轴上的线性关系,在局域、区域和大陆尺度上存在有趣的差异。
5. 局域种群丰富度与物种地理范围的大小之间几乎非常普遍的存在种间正相关,以及研究较少但正在出现的相同形式的种内相关性。
6. 纬度与物种丰富度之间有非常紧密的联系,但也有偏离广泛格局的情况。
7. 生态系统的能量输入和物种丰富度之间的相关但不同的格局。
8. 在通常被认为是生态系统科学中心的地方,也有大量醒目的宏观格局。
9. 化石记录提供了关于生态过程的完全不同的观点。
7. 结束语
我可以继续说下去,但这些已经很足够了。这些以及其他宏观生态学格局,为生态学相互作用的大量细节设定了边界条件。这些格局表明了生态学的偶然性定律和规律框架。这些格局本身来自于支配我们学科的四个真正非常普遍的定律。从这四条非常普遍的定律的相互作用中,自然界创造了辉煌的生命多样性,研究起来非常有趣,而且惊喜不断。当系统不太复杂时,也就是当偶然性事件可控时(如单一物种的种群动态,或非常小数量的相互作用的物种),以及在非常大的尺度上,当一种统计秩序从混乱中出现时,普遍的生态学特征就会从这种辉煌的多样性中最明显地显现出来。中间地带是一片混乱。它是令人着迷的研究,并富含奇妙的生物学知识。但是研究它室,不要期望出现普适的规律,甚至是简单的偶然的普适性规律。如果它们出现了,请珍惜它们。
地址:浙江省丽水市学院路1号电话:0578-2171795Email:zglslsxy@lsu.edu.cn
中国(丽水)两山学院版权所有制作维护